URBAN METAGENOMICS
http://metasub.org/city-profiles/
http://www.pathomap.org/2015/02/05/gc-timecourse/
http://elizabeth-henaff.net/Microbiome-of-the-NYC-subway
http://urbaneco.washington.edu/wp/research/urban-microbiome/
https://www.thenatureofcities.com/invisible-city-life-the-urban-microbiome/
https://weill.cornell.edu/researchers-produce-first-map-of-new-york-city-subway-system-microbes-christopher-mason.html
https://dirt.asla.org/designing-cities-for-healthier-human-microbiomes/
Designing Cities for Healthier Microbiomes
by Jared Green / 06/25/2018
“Humans are essentially super-organisms or holobionts made up of both human cells and those of micro-organisms, such as viruses, bacteria, archea, protists, and fungi. Researchers now know the human body hosts a comprehensive ecosystem, largely established by age three, in which non-human cells vastly outnumber human cells.
The latest study from the American Academy of Microbiology estimates each human ecosystem contains around 100 trillion cells of micro-organisms and just 37 trillion human cells. But while rainforest or prairie ecosystems are now well-understood, the human ecosystem is less so. As researchers make new discoveries, there is a growing group of scientists who argue our microbiomes are deeply connected with our physical and mental health.
The increased number of prebiotics and probiotics supplements on the shelf in drug stores and supermarkets, and availability of fresh pickles and kimchi in local farmers markets, are perhaps testaments to this increasingly-widespread belief. The question at the Environmental Design Research Association (EDRA) conference in Oklahoma City was: Can we design cities to better support our microbiomes and in turn our overall health?
Richard Wener, an environmental psychologist at NYU, explained how our built environment is filled with micro-organisms. “The walls of our kitchen and bathrooms are covered in bacteria; most of them aren’t pathogenic.” The micro-organisms found in our environments are constantly interacting with our microbiomes, so we are “perpetually assembling and re-assembling different species.”
For Wener, this begs the question: “What is an individual?” If we are constantly evolving with the micro-organisms in our environment and therefore changing our composition, “what individual are we talking about?” While our microbiomes may evolve, they still have a distinct signature. Researchers can now identify people by their unique microbiotic markers — crime-scene microbes are now analyzed in forensic studies to identify who was at a location.
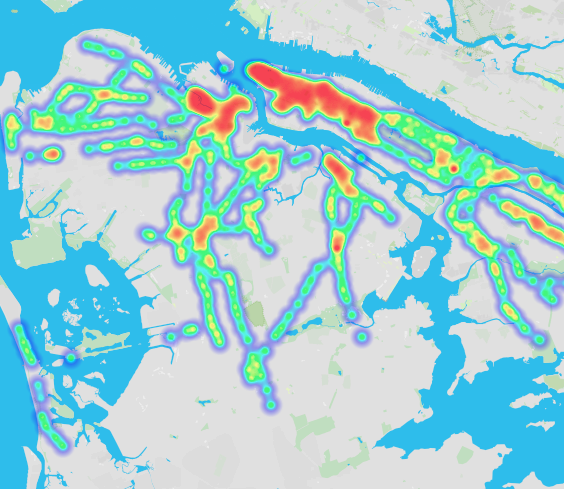
Places have unique microbiotic signatures as well. Through their PathoMap project, Weil Cornell Medical College organized citizen scientists to examine the bacteria in subway stations and found there were “consistent signatures around subway stations.” The microbes in subway stations near hospitals were different from neighborhood signatures. “The microbes really depend on who lives there.” (See data visualizations of the findings).
So can we design the city in terms of microbial species? Can we design buildings and parks to create “selective pressure that supports biotic life that is good for humans?” Turns out good microbes can be put to better use while bad ones can be suppressed.
In Brooklyn’s Gowanus Canal, a highly-polluted Super Fund site, researchers took samples of sludge at the bottom of the canal and found bacteria had been digesting some of the worst industrial waste for decades. “They co-evolved to eat solvents and toxins.” At a slow pace, Wener thinks these bacteria could actually clean-up the entire canal. Through interventions, “we could encourage the growth of this bacteria.”
Buildings could be designed to kill off pathogenic bacteria and support the healthy microbes in our biomes. Wener said a recent Brazilian study found pathogenic bacteria, which comes from people, thrives in closed spaces. In open air environments in the Brazilian rainforest, where lots of animals roamed through indigenous villages, there were no pathogenic bacteria, but in closed building interiors found in Brazilian cities, researchers found many. The conclusion is clear: If you increase exposure to light and air flow in buildings, you will reduce dangerous bacteria.
Through urban farming and gardening — or just plain playing in the dirt — humans can also increase their exposure to healthy microbes found in soils. A group of scientists and advocates argue that greater exposure could help fight depression and anxiety and reduce rates of asthma and allergies in both kids and adults.
https://youtu.be/G6WxEQrWUik
The incredible increase of allergies among Western populations may be caused by our “sterile, germ-free environments” that cause our immune systems to over-react to everything from nuts to mold and pollen. Dr. Brett Finlay and Marie-Claire Arrieta even wrote a book exploring this: Let Them Eat Dirt: Saving Your Child from an Over-sanitized World.
Wener said we have created cities that reflect our fear of bacteria; instead we must create microbial-inclusive cities that improve our health. “Most microbes in our bodies have co-evolved with us. They are important to our vital functions. The future of urban planning and design should support healthy microbes.”

As part of this vision, landscape architects could design parks and plazas to be filled with accessible garden plots and soil-based play areas that let both adults and kids get dirty. We could design for holobionts instead of just people, boosting the health of the collective urban microbiome in the process.

Wener’s colleage at NYU — Elizabeth Henaff — is leading much of this research. Learn about her artful experiments. Read this article from Michael Pollan in The New York Times outlining the connections between our microbiome and health, and this Q&A from The Guardian.”
ANCIENT THREATS
https://www.sciencemag.org/news/2018/06/momentous-transition-multicellular-life-may-not-have-been-so-hard-after-all
Transition to multicellular life may not be so hard after all
by Elizabeth Pennisi / Jun. 28, 2018
“Billions of years ago, life crossed a threshold. Single cells started to band together, and a world of formless, unicellular life was on course to evolve into the riot of shapes and functions of multicellular life today, from ants to pear trees to people. It’s a transition as momentous as any in the history of life, and until recently we had no idea how it happened.
The gulf between unicellular and multicellular life seems almost unbridgeable. A single cell’s existence is simple and limited. Like hermits, microbes need only be concerned with feeding themselves; neither coordination nor cooperation with others is necessary, though some microbes occasionally join forces. In contrast, cells in a multicellular organism, from the four cells in some algae to the 37 trillion in a human, give up their independence to stick together tenaciously; they take on specialized functions, and they curtail their own reproduction for the greater good, growing only as much as they need to fulfill their functions. When they rebel, cancer can break out.
Multicellularity brings new capabilities. Animals, for example, gain mobility for seeking better habitat, eluding predators, and chasing down prey. Plants can probe deep into the soil for water and nutrients; they can also grow toward sunny spots to maximize photosynthesis. Fungi build massive reproductive structures to spread their spores. But for all of multicellularity’s benefits, says László Nagy, an evolutionary biologist at the Biological Research Centre of the Hungarian Academy of Sciences in Szeged, it has traditionally “been viewed as a major transition with large genetic hurdles to it.”

“To study developing multicellular animals, students apply various stains. In juvenile squid (Loligo pealei) they reveal muscle (red), hairlike cilia (green), and cell nuclei (blue).”
Now, Nagy and other researchers are learning it may not have been so difficult after all. The evidence comes from multiple directions. The evolutionary histories of some groups of organisms record repeated transitions from single-celled to multicellular forms, suggesting the hurdles could not have been so high. Genetic comparisons between simple multicellular organisms and their single-celled relatives have revealed that much of the molecular equipment needed for cells to band together and coordinate their activities may have been in place well before multicellularity evolved. And clever experiments have shown that in the test tube, single-celled life can evolve the beginnings of multicellularity in just a few hundred generations—an evolutionary instant.
Evolutionary biologists still debate what drove simple aggregates of cells to become more and more complex, leading to the wondrous diversity of life today. But embarking on that road no longer seems so daunting. “We are beginning to get a sense of how it might have occurred,” says Ben Kerr, an evolutionary biologist at the University of Washington in Seattle. “You take what seems to be a major step in evolution and make it a series of minor steps.”
Hints of multicellularity date back 3 billion years, when impressions of what seem to be mats of microbes appear in the fossil record. Some have argued that 2-billion-year-old, coil-shaped fossils of what may be blue-green or green algae—found in the United States and Asia and dubbed Grypania spiralis—or 2.5-billion-year-old microscopic filaments recorded in South Africa represent the first true evidence of multicellular life. Other kinds of complex organisms don’t show up until much later in the fossil record. Sponges, considered by many to be the most primitive living animal, may date back to 750 million years ago, but many researchers consider a group of frondlike creatures called the Ediacarans, common about 570 million years ago, to be the first definitive animal fossils. Likewise, fossil spores suggest multicellular plants evolved from algae at least 470 million years ago.

“A slipper limpet (Crepidula fornicate) larva has a shell (green) and a membrane edged by a C-shaped line of nuclei (blue). Yellow shows nerves and purple a muscle that pulls the animal into its shell.”
Plants and animals each made the leap to multicellularity just once. But in other groups, the transition took place again and again. Fungi likely evolved complex multicellularity in the form of fruiting bodies—think mushrooms—on about a dozen separate occasions, Nagy concluded in a preprint posted 8 December 2017 on bioRxiv, based on a review of how different species of fungi—some single-celled, some multicellular—are related to one another. The same goes for algae: Red, brown, and green algae all evolved their own multicellular forms over the past billion years or so.
Nicole King, a biologist at the University of California (UC), Berkeley, found a revealing window on those ancient transitions: choanoflagellates, a group of living protists that seems on the cusp of making the leap to multicellularity. These single-celled cousins of animals, endowed with a whiplike flagellum and a collar of shorter hairs, resemble the food-filtering “collar” cells that line the channels of sponges. Some choanoflagellates themselves can form spherical colonies. More than 2 decades ago, King learned to culture and study these aquatic creatures, and by 2001 her genetic analyses were starting to raise doubts about the then-current view that the transition to multicellularity was a major genetic leap.
Her lab began turning up gene after gene once thought to be exclusive to complex animals—and seemingly unneeded in a solitary cell. Choanoflagellates have genes for tyrosine kinases, enzymes that, in complex animals, help control the functions of specialized cells, such as insulin secretion in the pancreas. They have cell growth regulators such as p53, a gene notorious for its link to cancer in humans. They even have genes for cadherins and C-type lectins, proteins that help cells stick together, keeping a tissue intact.

“Blood vessels (green), nuclei (blue), and actively dividing cells (red) color this 10.5-day-old mouse embryo, whose organs and body parts have already begun to emerge.”
All told, by surveying the active genes in 21 choanoflagellate species, King’s group found that these “simple” organisms have some 350 gene families once thought to be exclusive to multicellular animals, they reported on 31 May in eLife. If, as she and others believe, choanoflagellates offer a glimpse of the one-celled ancestor of animals, that organism was already well-equipped for multicellular life. King and her lab “have put protists at the front of research to address animal origins,” says Iñaki Ruiz-Trillo, an evolutionary biologist at the Spanish National Research Council and Pompeu Fabra University in Barcelona, Spain.
The ancestral versions of those genes might not have done the same jobs they later took on. For example, choanoflagellates have genes for proteins crucial to neurons, and yet their cells don’t resemble nerve cells, King says. Likewise, their flagellum has a protein that in vertebrates helps create the body’s left-right asymmetry, but what it does in the single-celled organism is unknown. And choanoflagellate genomes don’t anticipate multicellularity in every respect; they lack some critical genes, including transcription factors such as Pax and Sox, important in animal development. The missing genes give us “a better idea of what the actual animal innovations were,” King says.
As cells banded together, they didn’t just put existing genes to new uses. Studies of Volvox, an alga that forms beautiful, flagellated green balls, shows that multicellular organisms also found new ways to use existing functions. Volvox and its relatives span the transition to multicellularity. Whereas Volvox individuals have 500 to 60,000 cells arranged in a hollow sphere, some relatives, such as the Gonium species, have as few as four to 16 cells; others are completely unicellular. By comparing biology and genetics along the continuum from one cell to thousands, biologists are gleaning the requirements for becoming ever more complex. “What this group of algae has taught us is some of the steps involved in the evolution of a multicellular organism,” says Matthew Herron, an evolutionary biologist at the Georgia Institute of Technology in Atlanta.

“In this amphipod, Parhyale hawaiensis, muscle (pink), the circulatory system (yellow), and the hard covering called the cuticle (blue) work together to give this tiny arthropod its characteristic form and function.”
These studies show that many functions of specialized cells in a complex organism are not new. Instead, features and functions seen in single-celled organisms are rearranged in time and space in their multicellular relatives, says Corina Tarnita, a theoretical biologist at Princeton University. For example, in a unicellular relative of Volvox, Chlamydomonas, organelles called centrioles do double duty. For much of the cell’s lifetime they anchor the two whirling flagella that propel the cell through the water.
But when that cell prepares to reproduce, it loses the flagella, and the centrioles move toward the nucleus, where they help pull apart the dividing cell’s chromosomes. Later, the daughter cells each regrow the flagella. Chlamydomonas can both swim and reproduce, but not at the same time. Multicellular Volvox can do both at once, because its cells have specialized. The smaller cells always have flagella, which sweep nutrients over the Volvox‘s surface and help it swim. Larger cells lack flagella and instead use the centrioles full time for cell division.
Volvox has repurposed other features of the single cell ancestor as well. In Chlamydomonas, an ancient stress response pathway blocks reproduction at night, when photosynthesis shuts down and resources are scarcer. But in Volvox, the same pathway is active all the time in its swimming cells, to keep their reproduction permanently at bay. What was a response to an environmental signal in the single cell ancestor has been co-opted for promoting division of labor in its more complex descendent, Kerr says.

“This juvenile veiled chameleon (Chamaeleo calyptratus), with its bones in red and its cartilage in blue, reveals the skeleton’s main cell types.”
A third set of organisms hints at how this repurposing of existing genes and functions could have taken place. Over the past decade, Ruiz-Trillo and his colleagues have compared more than a dozen protist genomes with those of animals—a comparison that underscored the greater size and complexity of the animal genomes, they reported on 20 July in eLife. But a more telling finding came when Ruiz-Trillo; Arnau Sebé-Pedrós, now at the Weizmann Institute of Science in Rehovot, Israel; and Luciano di Croce at Barcelona’s Centre for Genomic Regulation analyzed the protist Capsaspora‘s portfolio of gene-regulating signals.
They found that the protist uses some of the same molecules as animals to turn genes on and off at particular times and places: proteins called transcription factors and long strands of RNA that don’t encode proteins. But its promoters—the regulatory DNA that interacts with transcription factors—were much shorter and simpler than in animals, the groups reported on 19 May 2016 in Cell, suggesting less sophisticated regulation. To Ruiz-Trillo and his team, the finding points to a key to multicellularity: increased fine-tuning of gene regulation.
What seemed a vast leap from single-celled ancestors looks less daunting if it was partly a matter of resetting the genetic switches, enabling existing genes to be active at new times and places. “This is what evolution always does, makes use of things that are around for new purposes,” says William Ratcliff of Georgia Tech. That thrifty repurposing may explain the swift transitions that have unfolded in Ratcliff ‘s lab. Instead of looking at the fossil record or comparing genomes of existing organisms, he has recreated evolution in lab cultures. “My own research has been not try to find out what happened in the real world, but to look at the process of how cells evolve increased complexity,” he explains.
“A slipper limpet (Crepidula fornicate) larva has a shell (green) and a membrane edged by a C-shaped line of nuclei (blue). Yellow shows nerves and purple a muscle that pulls the animal into its shell.”
As a postdoc working with Michael Travisano at the University of Minnesota in St. Paul, Ratcliff subjected yeast cultures to a form of artificial selection. He allowed only the biggest cells—measured by how fast they settled to the bottom of the flask—to survive and reproduce. Within 2 months, multicellular clusters began to appear, as newly formed daughter cells stuck to their mothers and formed branching structures.
As each culture continued to evolve—some have now been through more than 3000 generations—the snowflakes got bigger, the yeast cells became more durable and more elongated, and a new mode of reproduction evolved. In large snowflake yeast, a few cells along long branches undergo a form of suicide, releasing the cells at the tip to start a new snowflake. The dying cell sacrifices its life so that the group can reproduce. It’s a rudimentary form of cell differentiation, Ratcliff explains. He has just started to explore the genetic basis of these fast appearing traits; it appears to be a mix of existing genes being co-opted for new functions and other genes—such as one that helps dividing yeast cells separate—becoming disabled.
The yeast also developed a safeguard that is key to multicellularity: a way to keep cellular cheaters at bay. Such cheaters arise when mutations make some cells different from others, and possibly less cooperative. In complex organisms such as humans, protection comes in part from having an immune system to destroy aberrant cells. It also depends on a bottleneck between generations, in which a single cell (a fertilized egg, for example) serves as the starting point for the next generation. The result is that all cells in the new generation start out genetically identical. Snowflake yeasts have their own way of purging themselves of deviant cells. Because mutations accumulate over time, the most aberrant cells are found at the tips of the snowflakes. But they break off to form new colonies before they have a chance to become cheaters.

“This developing dwarf cuttlefish (Sepia bandensis) has nerve cells (faint red) not just in the brain, but also in the arms, tentacles, and mantle, where a gas-filled “cuttlebone” (purple) is located. Muscle and brain are green; eyes, yellow; and nuclei, blue.”
This mechanism also enables group traits to evolve in the yeast. Mutations in the cells released from each snowflake branch are passed on to all cells in the next colony. Consequently, subsequent snowflakes start out with new group traits—in the size and number of cells or the frequency and locations of suicide cells, for example—that become grist for further evolution. From that point on, it’s the assemblage, not individual cells, that’s adapting.
The yeast results weren’t a fluke. In 2014, Ratcliff and his colleagues applied the same kind of selection for larger cells to Chlamydomonas, the single-celled alga, and again saw colonies quickly emerge. To address criticism that his artificial selection technique was too contrived, he and Herron then repeated the Chlamydomonasexperiment with a more natural selective pressure: a population of paramecia that eat Chlamydomonas—and tend to pick off the smaller cells. Again a kind of multicellularity was quick to appear: Within 750 generations—about a year—two of five experimental populations had started to form and reproduce as groups, the team wrote on 12 January in a preprint on bioRxiv.
If multicellularity comes so easy, why did it take several billion years after the origin of life for complex organisms to become firmly established? Traditionally, researchers have blamed the early atmosphere’s low oxygen levels: To get enough oxygen, organisms needed the highest possible ratio of surface to volume, which forced them to stay small. Only after oxygen levels rose about 1 billion years ago could larger, multicellular organisms arise.

“Comparing Volvox, an alga with hundreds of cells (bottom), with its simpler relatives—the single-celled Chlamydomonas (top left) and the 4-to-16 cell Gonium (top right)—has revealed steps toward multicellularity.”
In 2015, however, Nicholas Butterfield, a paleontologist at the University of Cambridge in the United Kingdom, proposed that low oxygen levels actually favored the evolution of multicellularity in ancient marine organisms. Larger, multicellular organisms—with multiple flagella—were better at sweeping water past their cell membranes to harvest oxygen. Scarce nutrients in the ancient seas would have helped drive the next step, the evolution of specialized cell types, because more complex organisms can harvest food more efficiently. As for why complex organisms took so long to emerge, Butterfield thinks the lag reflects the time it took to evolve the more sophisticated gene regulation needed for multicellularity. Butterfield’s theory “is really quite elegant and simple, building on first principles of physics and chemistry, set into a deep geochemical, biogeochemical, and biophysical context,” says Richard Grosberg, an evolutionary biologist at UC Davis.
Once organisms had crossed the threshold to multicellularity, they rarely turned back. In many lineages, the number of types of cells and organs continued to grow, and they developed ever-more-sophisticated ways to coordinate their activities. Ratcliff and Eric Libby, a theoretical biologist at Umeå University in Sweden, proposed 4 years ago that a ratcheting effect took over, driving an inexorable increase in complexity. The more specialized and dependent on one another the cells of complex organisms became, the harder it was to revert to a single-cell lifestyle. Evolutionary biologists Guy Cooper and Stuart West at the University of Oxford in the United Kingdom recently confirmed that picture in mathematical simulations. “Division of labor is not a consequence but a driver” of more complex organisms, Cooper and West wrote on 28 May in Nature Ecology & Evolution. Touched off by the initial transition from one cell to many, a cycle of increasing complexity took hold, and the richness of multicellular life today is the result.
A decade ago, a radical theory of cancer emerged: that this plague of multicellular organisms arises when their cells rewind the evolutionary clock and revert to acting like unicellular life. Recently, David Goode, a computational cancer biologist at the Peter MacCallum Cancer Centre in Melbourne, Australia, and colleagues have found evidence to support that idea. They examined gene expression in seven types of solid tumors—including breast, stomach, and liver cancers—and traced the ancestry of the active genes they found. Genes that date all the way back to early single-celled eukaryotic organisms were revved up, Goode’s team reported last year in the Proceedings of the National Academy of Sciences. In contrast, genes unique to many-celled animals had gone quiet.
When an organism makes the leap to multicellularity, it must evolve gene regulatory networks to ensure its cells stop dividing at the appropriate time and function in step with their neighbors. Goode and his colleagues suggested that in cancer, mutations that cripple the networks cause those constraints to break down, giving genes suited to a unicellular lifestyle free rein to drive growth. Cancers seem to “undo the molecular constraints and controls that evolved to enable multicellular life,” Goode says.
“The idea that cancer represents a release of ancient genes from multicellularity is very appealing,” says Mark Vincent, a medical oncologist at the London Health Sciences Centre in Canada. “It explains a lot about what is otherwise mysterious about cancer,” such as how drug resistance arises, he says, explaining that “a lot of the treatments we have mimic ancient threats, which eukaryotic cells had to have survived.”
